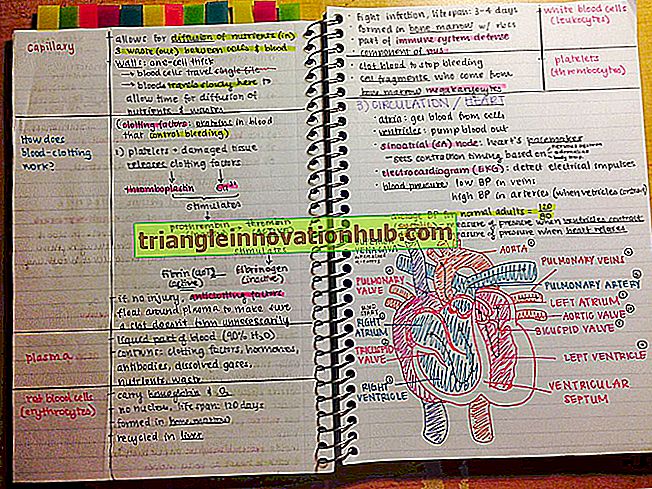
Processo de Metabolismo Secundário em Plantas
Processo de Metabolismo Secundário em Plantas!
As plantas não possuem nenhum mecanismo para o descarte de seus resíduos metabólicos. Esses resíduos se acumulam em suas células e muitas vezes servem como pontos de partida para muitos processos de transformação.
Tais vias, produzindo substâncias sem significado para a manutenção de uma via metabólica principal, constituem o que veio a ser chamado de metabolismo secundário.
O metabolismo secundário leva à formação de produtos secundários. As vias metabólicas secundárias são geneticamente e enzimaticamente reguladas. Os produtos secundários ou metabólitos secundários incluem alguns polissacarídeos, muitos antibióticos, esteróides, carotenóides, flavonóides alcalóides, etc.
Três tipos de produtos secundários são distinguidos em relação à estabilidade metabólica:
(1) produtos finais verdadeiramente inertes metabolicamente,
(2) produto cuja estabilidade se refere a um estado fisiológico ou de desenvolvimento e
(3) aqueles com um volume de negócios contínuo. No curso do metabolismo destes produtos, vários produtos poliméricos são formados: por exemplo, borracha, esporopollenins, lignins.
A formação destas substâncias é catalisada por fenoloxidases e peroxidases e, muitas vezes, a parede celular é o local preferido de formação destas macromoléculas. A degradação de produtos secundários a substratos metabólicos primários é efetuada por um grupo de enzimas que têm baixa especificidade de substrato, tais como monoxigenases e glicosidases metilantes.
As quantidades de produtos secundários variam com o estado de desenvolvimento da planta, a estação, o clima e até mesmo a hora do dia. Isso ocorre porque o metabolismo secundário está sujeito à influência de vários fatores reguladores.
O significado da formação de produtos secundários reside não tanto no fato de que eles servem como fonte de material para a regeneração de compostos necessários para o metabolismo primário. Mesmo que isso aconteça, a contribuição é tão pequena que não afeta o metabolismo como um todo.
Mas a formação de produtos secundários permite a continuação do evento metabólico primário, através da remoção de produtos da via sintética em causa do local de reação e isto é de muito maior importância.
Produtos secundários: Significado ecológico:
Os produtos secundários são de especial importância na manutenção do equilíbrio ecológico. A influência mútua entre os membros constituintes de um ecossistema é amplamente rastreável aos produtos secundários gerados pelas espécies componentes do sistema.
As relações alelopáticas entre uma planta e outra são, em certa medida, determinadas pela natureza dos produtos secundários. Por exemplo, os monoterpenos liberados das folhas de Salvia teucophylla e Artemisia californica tornariam o solo que absorvia aquelas substâncias impróprias para a geração das sementes das plantas anuais.
Produtos secundários: Relações planta-animal:
Os produtos vegetais secundários têm uma influência determinante sobre as relações planta-animal. Eles servem aos fins de predação, polinização e dispersão de sementes. O perfume desempenha um papel importante na atração dos insetos envolvidos com a polinização.
Substâncias envolvidas incluem monoterpenos como limoneno de citrus e geraniol de rosas; aminas como as monoaminas de Arum e derivados de ido benzco tais como vanilina de Baunilha. A cor da flor é a expressão de pigmentos, que também são produtos secundários.
Por exemplo, a cor branca de determinada flor é por causa da presença de flavonas como as luteolinas; flores amarelas são causadas por carotenóides, betaxantinas e flavonóis amarelos e as cores vermelha e violeta são causadas por betacianinas e antocianinas. Quando maduras, as frutas têm casacos de cores brilhantes e emitem odores característicos.
Ambos desempenham um papel significativo na atração de animais e aves e eventual dispersão de frutos e sementes. A cor e o odor originam-se em produtos secundários. Muitos produtos vegetais têm seu sabor e palatabilidade determinados pela presença de produtos secundários. Isso desempenha um papel em atrair ou repelir herbívoros.
Produtos secundários: Resistência a doenças e incidência:
Os produtos secundários fazem parte do sistema de defesa geral das plantas superiores contra infecções microbianas. Por outro lado, os parasitas formam produtos secundários que ajudam a superar a resistência da planta hospedeira.
Produtos secundários em relação ao homem:
Gosto, sabor, cor e palatabilidade dos alimentos e bebidas é devido à natureza química dos produtos secundários. Muitas das drogas que o homem usa também são produtos do metabolismo secundário nas plantas.
Por exemplo, D. Sorbital é um álcool de açúcar usado por diabéticos; a reserpina, uma droga anti-hipersensível, a estricnina, que serve como estimulante central, e a quinina, que é uma droga antimalárica, são todos derivados da Indole. Narcóticos como o haxixe e a maconha são monoterpenos da Cannabis sativa.
As bebidas de café, chá, cacau contêm cafeína, teobromina e teofilina, todas purinas. A nicotina do tabaco e a arecaidina da Areca catechu são derivados do ácido nicotínico. O valor da droga das plantas de Solanaceous como Datura e Atropa é devido à presença de escopolamina e outros alcalóides neles.
Uma grande variedade de produtos secundários serve como matéria-prima industrial. Óleo de linhaça empregado na indústria de tintas, óleo de mamona utilizado na fabricação de lubrificantes e sabões, ceras utilizadas na fabricação de vernizes e vernizes, suberina constituinte de cortiça utilizado na fabricação de isolantes e filtros são todos derivados de ácidos graxos.
Os taninos que têm uma aplicação importante na indústria do couro são os polifenóis. A borracha é um politerpeno produzido por Ficus elastica, Hevea brasiliensis, etc. Os óleos essenciais amplamente utilizados em perfumaria, cosméticos e preparações de solventes são terpenóides.
Mecanismo do Movimento Estomático:
Os estômatos funcionam como válvulas operadas por turgescência porque seu movimento de abertura e fechamento é governado por mudanças de turgor das células de proteção. Sempre que as células de proteção incham devido ao aumento do turgor, um poro é criado entre elas. Com a perda do turgor, os poros estomáticos são fechados. Os estômatos geralmente abrem durante o dia e fecham durante a noite com algumas exceções.
Os fatores importantes que governam a abertura estomática são: pH leve, alto ou reduzido e disponibilidade de água. Os fatores opostos governam o fechamento estomático, ou seja, escuridão, baixo pH ou alto CO 2 e desidratação. Existem três teorias principais sobre o mecanismo dos movimentos estomáticos:
1. Hipótese da Fotossíntese da Célula de Guarda (Schwendener, 1881):
As células de guarda contêm cloroplastos. Durante o dia, os cloroplastos realizam a fotossíntese e produzem açúcar. O açúcar aumenta a concentração osmótica de células-guarda. Causa a absorção de água das células epidérmicas próximas. As células de guarda túrgidas se curvam para fora e criam um poro no meio. No entanto, a atividade fotossintética dos cloroplastos das células-guarda parece ser insignificante.

2. Teoria da hidrólise do amido clássico:
As principais características da teoria foram explicitadas por Sayre (1923). Foi modificado por Steward (1964). As células guarda contêm amido. Na baixa concentração de dióxido de carbono (na manhã alcançada através da fotossíntese pelo mesofilo e células protetoras), o pH das células-guarda aumenta. Estimula a enzima fosforilase. A fosforilase converte o amido em glicose-1-fosfato. Este último é alterado para glicose-6-fosfato, que sofre hidrólise para produzir glicose e ácido fosfórico.

A glicose aumenta a concentração osmótica de células-guarda. Por causa disso, as células guarda absorvem água das células vizinhas, incham e criam um poro entre elas. O fechamento noturno dos estômatos é causado pelo aumento do conteúdo de dióxido de carbono (devido à parada da fotossíntese) da folha. Diminui o pH das células de guarda e provoca a fosforilação da glicose. Na presença de fosforilase, a glicose 1-fosfato é transformada em amido.

Como resultado, a concentração osmótica de células-guarda diminui. Eles perdem água para as células epidérmicas adjacentes. Com a perda de turgidez, as células-guarda encolhem e fecham o poro entre elas.
Objeções, (i) A glicose não é encontrada nas células-guarda no momento da abertura dos estômatos, (ii) Amido As mudanças de açúcar são quimicamente lentas, enquanto a abertura e o fechamento dos estômatos são bastante rápidos. (iii) Mudanças amplas no pH das células de proteção não podem ser explicadas com base na concentração de dióxido de carbono. (iv) Cebola e alguns de seus parentes não possuem amido ou polissacarídeo relacionado que pode ser hidrolisado até o nível de glicose; (v) A luz azul tem se mostrado mais efetiva que outros comprimentos de onda para abertura de estômatos. O mesmo não pode ser explicado pela teoria da hidrólise do amido, (vi) A hidrólise da teoria do amido não pode explicar o alto aumento na pressão osmótica encontrado nas células-guarda.
3. Hipótese da Bomba de Malato ou íon K + (Teoria Moderna):
As principais características da teoria foram apresentadas por Levitt (1974). De acordo com essa teoria, o pH da célula-guarda pode aumentar devido à absorção ativa de H + pelos cloroplastos das células-guarda ou mitocôndrias, assimilação de CO2 por mesofilo e células-guarda. Um aumento no pH provoca a hidrólise do amido para formar ácidos orgânicos, especialmente o piruvato de fosfoenol. Fosfato de Hexose de Amido -> Ácido Fosfoglicerico -> Piruvato de Fosfeno.
O piruvato de fosfoenol também pode ser formado pelo ácido pirúvico da via respiratória. Com a ajuda de PEP carboxylase (PEP case), combina com o CO2 disponível para produzir ácido oxálico que se transforma em ácido málico. O ácido málico dissocia-se em H + e malato.
Os íons H + passam para fora das células-guarda em troca de íons K + . Alguns íons K + (acompanhados por íons CI) também são absorvidos ativamente das células epidérmicas adjacentes com a ajuda de cAMP, ATP e citocininas. As células de guarda mantêm sua eletroneutralidade, equilibrando K + com malato e CI - .
No estado combinado, eles passam para os pequenos vacúolos e aumentam a concentração osmótica das células-guarda. Como resultado, as células de proteção absorvem a água das células epidérmicas próximas por meio da endosmose, incham e criam um poro entre elas.

Durante o fechamento dos estômatos, os íons H + se difundem dos cloroplastos das células-guarda. Diminui o pH do citoplasma da célula-guarda. Qualquer malato presente no citoplasma combina com H + para formar ácido málico. O excesso de ácido málico inibe sua própria biossíntese.
A alta concentração de CO 2 também tem um efeito similar. O ácido málico não dissociado promove o vazamento de íons (Jackson e Taylon, 1970). Como resultado, os íons K + se dissociam do malato e passam para fora das células-guarda. A formação de ácido abcísico (como durante a seca ou ao meio-dia) também promove a reversão da bomba de H + K + e aumenta a disponibilidade de H + dentro do citoplasma da célula-guarda.
A perda de íons K + diminui a concentração osmótica de células-guarda em comparação às células epidérmicas adjacentes. Isso causa exosmose e, consequentemente, a turgescência das células de proteção diminui. Fecha o poro entre as células de guarda. Simultaneamente, os ácidos orgânicos são metabolizados para produzir amido.